Endangered Species Recovery Program | |
|
Home | News | Publications | Species profiles | Data and maps | About | Staff | Links | Department of Biological Sciences | CSU Stanislaus |
Recovery Plan for Upland Species of the San Joaquin Valley, California
Contents
. Introduction
. Species accounts
. Recovery
. Stepdown
. Implementation
. References
. Appendix
A. California Jewelflower (Caulanthus californicus)
- 1. Description and Taxonomy
- 2. Historical and Current Distribution
- 3. Life History and Habitat
- 4. Reasons for Decline and Threats to Survival
- 5. Conservation Efforts
- 6. Recovery Strategy
1. Description and Taxonomy
Taxonomy.-- The appropriate genus for California jewelflower has been debated (Payson 1923, Rollins 1971, Al-Shehbaz 1973), but it has been recognized consistently as a distinct species. Watson (1880) originally named the genus Stanfordia solely to accommodate this species. The type specimen (i.e., the individual plant on which the original description was based) of Stanfordia californica was collected "near Tulare" in Tulare County. Greene then transferred the species to Streptanthus in 1891 (Greene 1891 as cited in Taylor and Davilla 1986). The currently accepted scientific name for California jewelflower, Caulanthus californicus, was published by Payson (1923). California jewelflower is a member of the mustard family (Brassicaceae).
Description.-- California jewelflower has hairless, usually branching stems, which can range from less than 10 centimeters (4 inches) to more than 50 centimeters (20 inches) tall (Munz and Keck 1959, Mazer and Hendrickson 1993a, Cypher 1994a). The upper leaves are egg-shaped and clasp the stem, unlike the leaves at the base of the plant, which are oblong. The maroon buds are clustered at the tip of the stem and contrast with the translucent, white flowers below. The fruits of California jewelflower are 1 to 6 centimeters (0.4 to 2.4 inches) long, and flattened (Buck 1993).
Identification.-- California jewelflower (Figure 5) differs from all other species of Caulanthus in that it has flattened, sword-shaped fruits and spherical seeds. Other jewelflowers also have maroon buds and whitish flowers, but those that overlap in range with California jewelflower have narrow, elongated fruits and flattened seeds (Buck 1993).
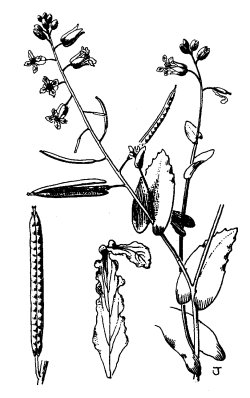
Figure 5. Illustration of California jewelflower (from Abrams, Vol. 2, 1944, with permission).
2. Historical and Current Distribution
Historical Distribution.-- The historical distribution of California jewelflower is known from 40 herbarium specimens, which were collected in 7 counties between 1880 and 1973. Approximately half of the collection sites were on the floor of the San Joaquin Valley in Fresno, Kern, and Tulare Counties (Figure 6). Several other collections came from two smaller valleys southwest of the San Joaquin Valley: the Carrizo Plain (San Luis Obispo County) and the Cuyama Valley (Santa Barbara and Ventura Counties). Three occurrences (i.e., collection sites separated by 0.4 kilometer [0.25 mile] or more) were in the Sierra Nevada foothills at the eastern margin of the San Joaquin Valley in Kern County. The remainder of the historical sites were in foothills west of the San Joaquin Valley, in Fresno, Kern, and Kings Counties (CDFG 1995, Taylor and Davilla 198).
Current Distribution.-- By 1986, all the occurrences on the San Joaquin and Cuyama Valley floors had been eliminated, and the only natural population known to be extant (i.e., still in existence) was in Santa Barbara Canyon, which is adjacent to the Cuyama Valley in Santa Barbara County (Taylor and Davilla 1986). A small, introduced colony also existed at the Paine Preserve in Kern County at that time. Since then, several more introductions have been attempted (see Conservation Efforts), and a number of colonies were rediscovered in two other areas where the species had been collected historically. The naturally-occurring populations of California jewelflower that are known to be extant today are in three centers of concentration: (1) Santa Barbara Canyon, (2) the Carrizo Plain in San Luis Obispo County, and (3) the Kreyenhagen Hills in Fresno County (CDFG 1995, Danielsen et al. 1994, B. Delgado pers. comm., R. Lewis pers. comm.).
The Santa Barbara Canyon metapopulation (i.e., scattered groups of plants that may function as a single population due to occasional interbreeding) occurs in an area of approximately 10 by 1 kilometer (6.5 by 0.5 mile) on the terraces just west of the Cuyama River and includes approximately 12 hectares (30 acres) of occupied habitat. The Carrizo Plain metapopulation is confined to the western side of the Carrizo Plain in a roughly triangular area measuring approximately 15 by 13 by 10 kilometers (9 by 8 by 6 miles) and encompassing approximately 4 hectares (10 acres) of occupied habitat (R. Lewis pers. comm.). The Kreyenhagen Hills metapopulation includes 4 small colonies within a 3 by 1 kilometer (2 by 0.5 mile) area of rolling hills (USBLM in litt. 1994).
Additional populations of California jewelflower may persist in the foothills of Fresno, Kern, and Kings Counties, where potential habitat remains in rangeland. However, access to historical sites in these areas has been restricted, so the presence of the species has not been verified in over 50 years (CDFG 1995, Taylor and Davilla 1986).
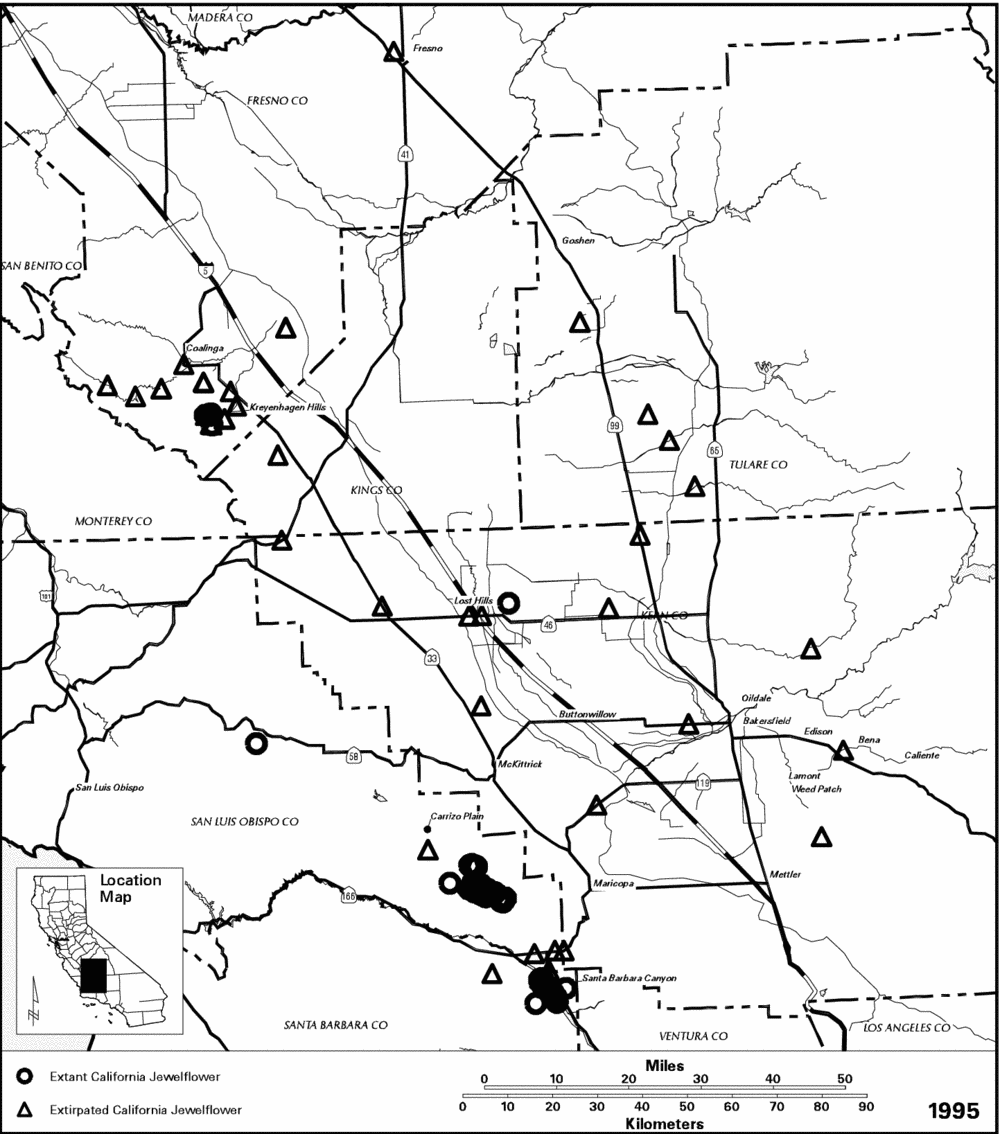
Figure 6. Distribution of California jewelflower (Caulanthus californicus).
3. Life History and Habitat
California jewelflower is an annual, meaning that each plant lives less than 1 year, and the entire life cycle from seed germination to seed set is completed in a single growing season. As is typical of annuals, both plant size and population size in California jewelflower can vary dramatically, depending on site and weather conditions (Taylor and Davilla 1986, Mazer and Hendrickson 1993a, Cypher 1994a).
Reproduction and Demography.-- Seeds of California jewelflower begin to germinate in the fall when the rainy season begins, but additional seedlings may continue to emerge for several months. California jewelflower seedlings develop into rosettes (clusters of leaves at ground level) during the winter months, and the stem elongates as flower buds begin to appear in February or March. Flowering and seed set continue until the plants die, which may occur as late as May in years of favorable rainfall and temperatures. Seed-dispersal agents are not known, but those that have been suggested for California jewelflower and related genera include gravity, seed-eating animals (Cypher 1994a), wind, and water (Al-Shehbaz 1973).
California jewelflower probably forms a persistent seed bank. In greenhouse trials, viable seeds did not germinate even when exposed to a variety of typical temperature and moisture condiions (Taylor and Davilla 1986). Only conditions simulating prolonged weathering induced seed germination (Mazer and Hendrickson 1993a). A persistent seed bank ensures that some seeds will be available to produce plants in succeeding years, even if no individuals survive to set seed in one unfavorable growing season (Philippi 1993). The presence of a seed bank would explain the reappearance of California jewelflower in uncultivated areas where it had not been observed for decades.
Pollinator-exclusion experiments indicated that insects are necessary for seed set in California jewelflower (Mazer and Hendrickson 1993a). Honeybees (Apis mellifera) have been observed visiting the flowers (R. Lewis pers. comm.), but native insects also would be expected to serve as pollinators. Solitary bees of the genus Synhalonia are known to visit Coulters jewelflower (Caulanthus couteri) (Thorp in litt. 1998). Closely-related species of the genus Thelypodium were visited by several species of bees (Bombus sp., Apis sp., and Xylocopa sp.) and butterflies (Pieris sp.) (Al-Shehbaz 1973). Both cross- and self-pollination resulted in seed set in greenhouse trials (Mazer and Hendrickson 1993a).
In 1992 and 1993, which were years of above-average rainfall during the growing season, 46 percent to 85 percent of plants in study areas on the Carrizo Plain survived long enough to produce seed (Mazer and Hendrickson 1993a, Cypher 1994a). However, in years of below-average precipitation or above-average temperatures, all the plants may die before setting seed (R. van de Hoek pers. comm.). Seed production in California jewelflower may vary greatly among individuals, sites, and years. In 1992, average seed production per plant was 711 on the Carrizo Plain and 278 in Santa Barbara Canyon (Mazer and Hendrickson 1993a). In 1993, the estimated number of seeds per plant on the Carrizo Plain ranged from 4 to over 11,000 and averaged 929, compared to 49 in the Kreyenhagen Hills (E. Cypher unpubl. data).
Habitat and Community Associations.-- Extant populations of California jewelflower occur in Nonnative Grassland, Upper Sonoran Subshrub Scrub, and Cismontane Juniper Woodland and Scrub (E. Cypher unpubl. data). Historical records suggest that California jewelflower also occurred in the Valley Saltbush Scrub community in the past (CDFG 1995).
Herbaceous cover was dense at most California jewelflower sites studied in 1993 (Cypher 1994a). Native plant species, such as annual fescue (Vulpia microstachys), clovers (Trifolium spp.), red maids (Calandrinia ciliata), and goldfields (Lasthenia californica) comprised a high proportion of the vegetation at many of the known locations over several years. The exotic grass red brome (Bromus madritensis ssp. rubens) was a significant component of the vegetation only at the Carrizo Plain sites (Taylor and Davilla 1986, Lewis in litt. 1993, Cypher 1994a, E. Cypher unpubl. data). On the Carrizo Plain, California jewelflower occurred primarily on the burrow systems of giant kangaroo rats (Dipodomys ingens), another endangered species (Cypher 1994a).
Populations of California jewelflower have been reported from elevations of approximately 75 to 900 meters (240 to 2,950 feet) and from level terrain to 25 percent slopes. Soils at known sites are primarily subalkaline, sandy loams (CDFG 1995, Taylor and Davilla 1986, Lewis in litt. 1993).
4. Reasons for Decline and Threats to Survival
Reasons for Decline.-- The primary reason for the decline o California jewelflower was habitat destruction. All the populations on the San Joaquin and Cuyama Valley floors have been eliminated. Conversion to agriculture accounted for the loss of most sites, but those closest to Bakersfield and Fresno were destroyed by urbanization. Oilfield activity may have eliminated a few sites in the foothills at the western margin of the San Joaquin Valley (Taylor and Davilla 1986).
Threats to Survival.-- Development remains a threat in Santa Barbara Canyon, where more than 90 percent of the California jewelflower metapopulation occurs on private land. The California jewelflower habitat near the canyon mouth is for sale; the landowner cleared California junipers (Juniperus californica) from the site and planted ornamentals in anticipation of residential development (Lewis in litt. 1993). California jewelflower on private land in the upper portion of Santa Barbara Canyon is subject to cattle grazing throughout the growing season, but the magnitude of threat posed by livestock is unknown. Grazing in the period between the rosette stage and seed set is believed to be detrimental to California jewelflower because seed set would be reduced if flowering or fruiting stems were consumed (Mazer and Hendrickson 1993a, R. Lewis pers. comm.).
Potential threats to one or more of the remaining populations of California jewelflower include competition from exotic plants, the effects of certain insecticides on pollinators, and small population size. In a preliminary study, seedling mortality was higher in plots that contained dense vegetation than in plots where all plants but California jewelflower had been removed (Mazer and Hendrickson 1993a). Red brome could be particularly competitive because some strains are resistant to air pollution (Westmann et al. 1985 in Taylor and Davilla 1986). Insecticides could present a threat to California jewelflower viability on the Carrizo Plain by decreasing pollinator populations. Prior to 1980, the California Department of Food and Agriculture sprayed malathion on the Carrizo Plain to control beet leafhoppers. The effect of malathion on native insect populations has not been investigated. Thus, it is unknown whether fall spraying would affect pollinator populations the following spring, or how large a buffer zone would be needed to avoid affecting insects that pollinate California jewelflower. Under the current environmental assessment and pesticide use permit, spraying has been suspended on the Carrizo Plain, at least through the year 2001 (California Department of Food and Agriculture in litt. 1998). Small population size may be another factor in the continued existence of California jewelflower.
5. Conservation Efforts
California jewelflower was state listed as endangered in 1987 and federally listed as endangered in 1990 (USFWS 1990; Table 1). Intensive and extensive survey efforts were undertaken by biologists from the California Energy Commission, USBLM, and U.S. Forest Service, beginning in 1987. Their efforts led to the discovery of many new occurrences and the rediscovery of several that were thought to have been eliminated. Surveys for additional populations are continuing in suitable habitats on the Los Padres National Forest and USBLM lands in the Bakersfield District (Danielsen et al. 1994, B. Delgado pers. comm., R. Lewis pers. comm.).
The known California jewelflower habitat in two of the three concentration areas is in public ownership. The Carrizo Plain metapopulation is entirely within the Carrizo Plain Natural Area, which is administered jointly by USBLM, The Nature Conservancy, and CDFG. USBLM also administers the Kreyenhagen Hills and a small part of Santa Barbara Canyon. Populations in each of these areas have been monitored annually by USBLM personnel since 1991. USBLM no longer allows green-season grazing in California jewelflowe habitats under its management, which include approximately 40 percent of individuals known to be extant. In 1994, an exclosure was constructed around the plants on public land in Santa Barbara Canyon to preclude grazing (R. Lewis pers. comm.).
Several experimental introductions of California jewelflower have been attempted in Kern, Santa Barbara, and Tulare Counties on lands protected by The Nature Conservancy and the Los Padres National Forest (Taylor 1988, CDFG 1995). In all instances, the number of plants at each site has declined precipitously following the initial seeding (Taylor and Davilla in litt. 1986, Danielsen et al. in litt. 1994). Possible causes of failure included unfavorable site conditions, use of seed sources that were not adapted to the introduction site, lack of genetic diversity in the introduced populations, and insufficient numbers of seeds (Taylor and Davilla 1986, Mazer and Hendrickson 1993a, Danielsen et al. 1994). Considering the variable germination rates in natural populations, plants may reappear at some of the reintroduction sites after several years. Future reintroduction efforts can build on the experience gained from these early trials.
Preliminary research on the reproductive biology, demography, and ecology of California jewelflower has been conducted by Dr. Susan Mazer and associates from the University of California, Santa Barbara, and by Dr. Ellen Cypher and associates from the Endangered Species Recovery Program. Funding for these studies was provided by CDFG, the National Science Foundation, USBLM, and Endangered Species Recovery Program. The U.S. Natural Resources Conservation Service is considering artificial propagation of California jewelflower to aid research and restoration efforts (D. Dyer pers. comm.).
6. Recovery Strategy
Although restoration of California jewelflower to all its former sites of occurrence is not feasible, the recovery goal is to maintain self-sustaining populations in protected areas representative of the former geographic and topographic range of the species and in a variety of appropriate natural communities. Surveys will be necessary to determine whether natural populations remain in all target areas. Where natural populations no longer exist, such as the floor of the San Joaquin Valley, reintroduction will be necessary to achieve recovery. However, reintroduction is expensive and experimental, and thus the preferable course of action is to locate and protect the remaining occupied habitat wherever possible. Unoccupied habitat within metapopulations also should be protected to facilitate movement of pollinators and seed dispersers. Thus, additional elements of the strategy are to protect land in blocks of at least 65 hectares (160 acres) and to avoid fragmenting any metapopulation into more than two blocks of contiguous, protected natural land. Finally, buffer zones of 150 meters (500 feet) or more should be protected beyond the population margins to reduce external influences and to allow for population expansion.
The top-priority action for recovery of California jewelflower is to protect the plants on private land in Santa Barbara Canyon. The site could be secured through fee title acquisition or conservation easements. Continued protection and appropriate management of all occupied habitat on public lands also is important. A number of additional tasks are required to achieve recovery goals. These tasks include developing management plans, surveying for additional populations, banking seed, conducting research, and modeling population demographics using matrix projection modeling. Interim management plans should be developed for each protected area to ensure that recovery of California jewelflower and other listed species is the primary goal. Management plans may need to be revised if populations begin declining or research identifies limitations to population viability.
Surveys are particularly important in the foothills east of the San Joaquin Valley to determine if historical populations remain extant. If populations are rediscovered in that area thir protection would be a high-priority task because they are likely to incorporate genotypes not found elsewhere in the range. Seeds should be collected from each of the known metapopulations and any occurrences discovered in the future, according to the guidelines established by the Center for Plant Conservation (1991). Seed collections should be used for two purposes: to conserve the genetic diversity of the species in seed-banking facilities; and to allow greenhouse propagation of the species, which would allow experimental introductions and manipulative studies without jeopardizing natural populations or continuing to deplete natural seed banks (Mazer and Hendrickson 1993a). Continued demographic research is necessary to determine which stages in the life cycle are limiting (Schemske et al. 1994). Limiting factors may vary among California jewelflower populations and can include pollinator availability, competition from introduced plants, consumption by kangaroo rats or livestock, or physical site characteristics (Mazer and Hendrickson 1993a, Cypher 1994a). Annual monitoring also is necessary to indicate whether population levels are increasing, decreasing, or remaining stable (Cypher 1994a, Schemske et al. 1994).